Лягушка - типичный представитель земноводных. На примере этого животного можно изучить характеристики всего класса. Данная статья подробно описывает внутреннее строение лягушки.
Начинается пищеварительная система ротоглоточной полостью. На её дне прикреплен язык, который лягушка использует для ловли насекомых. Благодаря необычному строению он способен с большой скоростью выбрасываться изо рта и приклеивать к себе жертву.
На нёбных костях, а также на нижней и верхней челюсти земноводного имеются небольшие зубы конической формы. Служат они не для пережёвывания, а прежде всего для удержания добычи в пасти. Это ещё одно сходство амфибии с рыбами. Секрет, выделяемый слюнными железами, увлажняет ротоглоточную полость и пищу. Это облегчает её проглатывание. Пищеварительных ферментов слюна лягушки не содержит.

Пищеварительный тракт лягушки начинается глоткой. Далее следует пищевод, а затем - желудок. За желудком находится двенадцатиперстная кишка, остальная часть кишечника уложена в виде петель. Заканчивается кишечник клоакой. У лягушек есть и пищеварительные железы - печень и поджелудочная.
Пойманная с помощью языка добыча оказывается в ротоглотке, а затем через глотку попадает по пищеводу в желудок. Клетки, расположенные на стенках желудка, выделяют соляную кислоту и пепсин, которые способствуют перевариванию пищи. Далее полупереваренная масса следует в двенадцатиперстную кишку, в которую также изливаются секреты поджелудочной железы и впадает желчный проток печени.
Постепенно двенадцатиперстная переходит в тонкую кишку, где и всасываются все полезные вещества. Остатки пищи, которая не переварилась, попадают в последний отдел кишечника - короткую и широкую прямую кишку, оканчивающуюся клоакой.
Внутреннее строение лягушки и её личинки отличается. Взрослые особи являются хищниками и питаются в основном насекомыми, а вот головастики - самые настоящие травоядные. На их челюстях расположены роговые пластинки, с помощью которых личинки соскребают мелкие водоросли вместе с живущими в них одноклеточными организмами.
Дыхательная система
Интересные особенности внутреннего строения лягушки касаются и дыхания. Дело в том, что наравне с легкими, огромную роль в процессе газообмена играет наполненная капиллярами кожа амфибии. Легкие представляют собой тонкостенные парные мешки с ячеистой внутренней поверхностью и разветвленной сетью кровеносных сосудов.

Как дышит лягушка? Амфибия использует клапаны, способные открывать и закрывать ноздри, и движения дна ротоглотки. Для того чтобы сделать вдох, ноздри открываются, а дно ротоглоточной полости опускается, и воздух оказывается в пасти лягушки. Чтобы он прошел в легкие, ноздри закрываются, а дно ротоглотки поднимается. Выдох производится благодаря опаданию легочных стенок и движениям брюшных мышц.
У самцов гортанную щель окружают специальные черпаловидные хрящи, на которые натянуты голосовые связки. Высокая громкость звука обеспечивается благодаря голосовым мешкам, которые образованы слизистой оболочкой ротоглотки.
Выделительная система
Внутреннее строение лягушки, вернее, её тоже весьма любопытно, так как продукты жизнедеятельности амфибии могут выводиться через легкие и кожу. Но все же большая их часть выделяется почками, которые расположены у крестцового позвонка. Сами почки представляют собой продолговатые тела, прилегающие к спине. В этих органах есть специальные клубочки, способные отфильтровывать из крови продукты распада.
Моча по мочеточникам выводится в мочевой пузырь, где накапливается. После наполнения мочевого пузыря мышцы у брюшной поверхности клоаки сокращаются и жидкость выбрасывается через клоаку наружу.
Кровеносная система
Внутреннее строение лягушки более сложное, чем у взрослой лягушки трехкамерное, состоящее из желудочка и двух предсердий. Из-за единственного желудочка артериальная и венозная кровь частично смешивается, два круга кровообращения разобщены не полностью. Артериальный конус, имеющий продольный спиральный клапан, отходит от желудочка и распределяет смешанную и артериальную кровь в разные сосуды.

В правом предсердии собирается смешанная кровь: венозная поступает от внутренних органов, а артериальная - от кожи. В левое предсердие от легких поступает артериальная кровь.
Сокращаются предсердия одновременно, и кровь из обоих попадает в единственный желудочек. Благодаря строению продольного клапана поступает в органы головы и мозг, смешанная - к органам и частям тела, а венозная - к коже и легким. Школьникам может быть тяжело понять внутреннее строение лягушки. Схема кровеносной системы амфибии поможет наглядно представить, как работает кровообращение.
Кровеносная система головастиков имеет только один круг кровообращения, одно предсердие и один желудочек, как у рыб.
Строение крови лягушки и человека отличается. имеют ядро, овальную форму, а у человека - двояковогнутая форма, ядро отсутствует.
Эндокринная система
В эндокринную систему лягушки входят щитовидная, половая и поджелудочная железы, надпочечники и гипофиз. Щитовидная железа продуцирует гормоны, необходимые для завершения метаморфоза и поддержания обмена веществ, половые железы отвечают за размножение. Поджелудочная железа участвует в переваривании пищи, надпочечники помогают регулировать обмен веществ. Гипофиз вырабатывает ряд гормонов, влияющих на развитие, рост и окраску животного.

Нервная система
Нервная система лягушки характеризуется невысокой степенью развития, она схожа по характеристикам с нервной системой рыб, но имеет более прогрессивные черты. Головной мозг подразделяется на 5 отделов: средний, промежуточный, передний, продолговатый мозг и мозжечок. Передний мозг хорошо развит и разделяется на два полушария, каждое из которых имеет боковой желудочек - специальную полость.
В связи с однообразными движениями и в общем малоподвижным образом жизни, мозжечок имеет небольшой размер. Продолговатый мозг крупнее. Всего из головного мозга лягушки выходит десять пар нервов.
Органы чувств
Значительные изменения в органах чувств земноводных связаны с выходом из водной среды на сушу. Они уже сложнее, чем у рыб, так как должны помогать ориентироваться и в воде, и на суше. У головастиков развиты органы боковой линии.

В слое эпидермиса скрыты болевые, осязательные и температурные рецепторы. Сосочки на языке, небе и челюстях выполняют функцию органов вкуса. Органы обоняния состоят из парных обонятельных мешков, которые открываются как наружными, так и внутренними ноздрями, в окружающую среду и ротоглоточную полость соответственно. В воде ноздри закрыты, органы обоняния не действуют.
В качестве органов слуха развито среднее ухо, в котором находится аппарат, усиливающий звуковые колебания благодаря барабанной перепонке.
Строение глаза лягушки сложное, ведь ей необходимо видеть и под водой, и на суше. Защищают глаза взрослых особей подвижные веки и мигательная перепонка. У головастиков век нет. Роговица лягушачьего глаза выпуклая, хрусталик - двояковыпуклый. Амфибии видят довольно далеко и обладают цветным зрением.
137. Рассмотрите рисунки. Напишите названия частей тела лягушки. Какие органы расположены на ее голове? Напишите их названия.
138. Изучите таблицу "Класс Земноводные. Строение лягушки". Рассмотрите рисунок. Напишите названия внутренних органов лягушки, обозначенных цифрами.
3. желудок
4. поджелудочная железа
5. семенник
7. мочевой пузырь
8. толстая кишка
9. тонкая кишка
10. желчный пузырь
11. печень
139. Опишите строение головного мозга земноводных.
Головной мозг у земноводных имеет более прогрессивные черты, более крупные размеры переднего мозга, полное разделение полушарий. Средний мозг относительно небольшой. Мозжечок мал, так как у амфибий однообразные движения. От головного мозга отходят 10 черепно-мозговых нервов.
Отделы головного мозга: передний, средний, мозжечок, продолговатый, промежуточный.
140. Изучите таблицу "Класс Земноводные. Строение лягушки". Рассмотрите рисунок. Подпишите названия частей скелета лягушки, обозначенных цифрами.

2. лопатка
4. предплечье
9. уростиль
10. позвоночник
141. Рассмотрите рисунок. Напишите названия пищеварительной системы лягушки, обозначенных цифрами. Как осуществляется процесс пишеварения у лягушки?

1. ротовое отверстие
2. пищевод
3. желудок
4. кишечник
Все земноводные питаются итолько подвижной пищей. На дне ротовой полости находится язык. При ловле добычи он выкидывается изо рта, к нему прикрепляется добыча. На верхней челюсти имеются зубы, служащие только для удерживания добычи. При глотании проталкиванию пищи в пищевод помогают глазные яблоки.
В ротоглотку открываются протоки слюнных желез. Из ротоглотки пища по пищеводу попадает в желудок, оттуда в 12-перстную кишку. Сюда открываются протоки печени и поджелудочной железы. Переваривание происходит в желудке и 12-перстной кишке. Тонкий кишечник переходит в прямую, которая образует расширение - клоаку.
142. Зарисуйте схему строения сердца лягушки. Какую кровь называют артериальной, а какую - венозной?
Артериальная кровь идет от легких, богата кислородом. Венозная - к легким.

143. Опишите процесс размножения и развития лягушки. Укажите сходство и различие в размножении земноводных и рыб.
Размножаются земноводные в небольших хорошо прогреваемых участках водоемов. Органы размножения у самцов - семенники, у самок - яичники. Оплодотворение наружное.
Развитие лягушки: яйцо - головастик в момент выклева - развитие плавниковых складок и наружных жабр - стадия максимального развития наружных жабр - стадия исчезновения наружных жабр - стадия появления задних конечностей - стадия расчленения и подвижности задних конечностей - стадия освобождения передних конечностей, метаморфоза ротового аппарата и начала резорбции хвоста - стадия выхода на сушу.
144. Заполните таблицу.
Строение и значение органов чувств лягушки.
Органы чувств лягушки Особенности строения Значение Глаза Есть верхние (кожистые) и нижние (прозрачные) подвижные веки, мигательная перепонка. Есть железа, секрет которой смачивает роговицу и предохраняет ее от высыхания. Роговица выпуклая. Хрусталик имеет форму двояковыпукло линзы. У многих развито цветное зрение Зрение Орган слуха Внутренне ухо, среднее ухо. Наружно слуховое отверстие закрывает барабанная перепонка, содиненная со слуховой косточкой - стремячом Слух Орган равновесия Внутренне ухо Ориентировка Орган обоняния Парные обонятельные мешки. Их стенки выстланы обонятельным эпителием. Открываются наружу ноздрями, а в ротоглотку - хаонами Восприятие запахов Орган осязания Кожа Восприятие раздражения Орган боковой линии Боковая линия у личинок Позволяет ощущать ток воды
Строение головного мозга костистых рыб
Головной мозг костистых рыб состоит из типичных для большинства позвоночных пяти отделов.
Ромбовидный мозг (rhombencephalon)
передним отделом заходит под мозжечок, а сзади без видимых границ переходит в спинной мозг. Чтобы рассмотреть передний отдел продолговатого мозга, необходимо отвернуть вперед тело мозжечка (у некоторых рыб мозжечок невелик и передний отдел продолговатого мозга хорошо виден). Крыша в этом отделе мозга представлена сосудистым сплетением. Под ним лежит большая расширенная на переднем конце и переходящая сзади в узкую медиальную щель, она является полостью Продолговатый мозг служит местом отхождения большинства головных нервов, а также проводящим путем, связывающим различные центры передних отделов головного мозга со спинным. Однако, слой белого вещества, покрывающий продолговатый мозг, у рыб довольно тонкий, так как туловище и хвост в значительной степени автономны - они осуществляют большую часть движений рефлекторно, не соотносясь с головным мозгом. В дне продолговатого мозга у рыб и хвостатых амфибий лежит пара гигантских маутнеровских клеток, связанных с акустико-латеральными центрами. Их толстые аксоны простираются вдоль всего спинного мозга. Локомоция у рыб осуществляется в основном за счёт ритмичных изгибов тела, которыми, по-видимому, управляют главным образом местные спинномозговые рефлексы. Однако общий контроль над этими движениями осуществляют маутнеровские клетки. В дне продолговатого мозга лежит дыхательный центр.
Рассматривая мозг снизу, можно различить места отхождения некоторых нервов. От латеральной стороны передней части продолговатого мозга отходят три круглых корешка. Первый, лежащий наиболее краниально, принадлежит V и VII нервам, средний корешок - только VII нерву, и, наконец, третий корешок, лежащий каудально, является VIII нервом. За ними, также от боковой поверхности продолговатого мозга, несколькими корешками отходят вместе IX и X пара. Остальные нервы тонки и обычно обрываются при препарировании.
Мозжечок (cerebellum) довольно хорошо развит, имеет округлую или вытянутую форму, он лежит над передней частью продолговатого мозга непосредственно за зрительными долями. Задним своим краем он прикрывает продолговатый мозг. Выступающая вверх часть является телом мозжечка (corpus cerebelli). Мозжечок является центром точной регуляции всех моторных иннервации, связанных с плаванием и схватыванием пищи.
Средний мозг (mesencephalon) - часть ствола головного мозга, пронизанная мозговым водопроводом. В его состав входят крупные, вытянутые продольно зрительные доли (они видны сверху).
Зрительные доли, или зрительная крыша (lobis opticus s. tectum opticus) - парные образования, отделенные друг от друга глубокой продольной бороздой. Зрительные доли являются первичными зрительными центрами, воспринимающими возбуждение. В них заканчиваются волокна зрительного нерва. У рыб этот отдел головного мозга имеет главенствующее значение, это центр, оказывающий основное влияние на деятельность организма. Серое вещество, покрывающее зрительные доли, имеет сложное слоистое строение, напоминающее строение коры мозжечка или полушарий
От вентральной поверхности зрительных долей отходят толстые зрительные нервы, перекрещивающиеся под поверхностью промежуточного мозга.
Если вскрыть зрительные доли среднего мозга, то можно увидеть, что в их полости от мозжечка отделяется складка, носящая название мозжечковой заслонки (valvule cerebellis). По бокам от нее в дне полости среднего мозга выделяются два бобовидных возвышения, называемых полулунными телами (tori semicircularis) и являющихся дополнительными центрами статоакустического органа.
Передний мозг (prosencephalon) менее развит, чем средний, он состоит из конечного и промежуточного мозга.
Части промежуточного мозга (diencephalon) лежат вокруг вертикально расположенной щели Боковые стенки желудочка - зрительные бугры или таламус (thalamus ) у рыб и амфибий имеют второстепенное значение (как координационные чувствительные и двигательные центры). Крыша третьего мозгового желудочка - надбугорье или эпиталамус - не содержит нейронов. В его составе находится переднее сосудистое сплетение (сосудистая покрышка третьего желудочка) и верхняя мозговая железа - эпифиз (epiphisis). Дно третьего мозгового желудочка - подбугорье или гипоталамус у рыб формирует парные вздутия - нижние доли (lobus inferior). Перед ними лежит нижняя мозговая железа - гипофиз (hypophisis). У многих рыб эта железа плотно входит в специальное углубление в дне черепа и обычно при препаровке обрывается; тогда хорошо видна воронка (infundibulum). перекрест зрительных нервов (chiasma nervorum opticorum).
у костистых рыб по сравнению с другими отделами мозга очень мал. Большинство рыб (кроме двоякодышащих и кистепёрых) отличает эвертированное (вывернутое) строение полушарий конечного мозга. Они как бы «вывернуты» вентро-латерально. Крыша переднего мозга не содержит нервных клеток, состоит из тонкой эпителиальной перепонки (pallium), которая при препаровке обычно снимается вместе с оболочкой мозга. В этом случае на препарате видно дно первого желудочка, разделенное глубокой продольной бороздой на два полосатых тела. Полосатые тела (corpora striatum1) состоят из двух отделов, что можно видеть при рассмотрении мозга сбоку. Фактически, эти массивные структуры содержат материал полосатых тел и коры довольно сложного строения.
Обонятельные луковицы (bulbus olfactorius) прилегают к переднему краю конечного мозга. От них вперед идут обонятельные нервы. У некоторых рыб (например, у трески) обонятельные луковицы вынесены далеко вперёд, в таком случае они соединяются с мозгом обонятельными трактами.
Черепномозговые нервы рыбы.
Всего от головного мозга рыбы отходит 10 пар нервов. В основном (и по наименованию и по функциям) они соответствуют нервам млекопитающих.
Строение головного мозга лягушки
Головной мозг лягушки, как и других амфибий, характеризуется следующими особенностями по сравнению с рыбами:
а) прогрессивным развитием головного мозга, выразившимся в обособлении парных полушарий продольной щелью и развитием в крыше мозга серого вещества древней коры (archipallium);
б) слабым развитием мозжечка;
в) слабой выраженностью изгибов головного мозга, благодаря чему промежуточный и средний отделы ясно видны сверху.

Ромбовидный мозг (rhombencephalon)
Продолговатый мозг (myelencephalon, medulla oblongata) , в который краниально переходит спинной мозг, отличается от последнего большей шириной и отхождением от его боковых поверхностей крупных корешков задних черепномозговых нервов. На дорсальной поверхности продолговатого мозга находится ромбовидная ямка (fossa rhomboidea), вмещающая четвёртый мозговой желудочек (ventriculus quartus). Сверху он прикрыт тонкой сосудистой покрышкой, которая удаляется вместе с мозговыми оболочками. По вентральной поверхности продолговатого мозга проходит вентральная щель - продолжение вентральной щели спинного мозга. В составе продолговатого мозга идут две пары тяжей (пучков волокон): нижняя пара, разделённая вентральной щелью - двигательные, верхняя пара - чувствительные. В продолговатом мозге расположены центры челюстного и подъязычного аппаратов, органа слуха, а также пищеварительной и дыхательной систем.
Мозжечок (cerebellum) располагается впереди ромбовидной ямки в виде высокого поперечного валика как выроста её передней стенки. Небольшие размеры мозжечка определяются небольшой и однообразной подвижностью амфибий - фактически, он состоит из двух небольших частей, тесно связанных с акустическими центрами продолговатого мозга (эти части сохраняются у млекопитающих как клочки мозжечка (flocculi)). Тело мозжечка - центр координации с другими отделами мозга - развито очень слабо.
Средний мозг (mesencephalon) при рассмотрении с дорсальной стороны представлен двумя типичными зрительными долями (lobus opticus s. tectum opticus) , имеющими вид парных яйцевидных возвышений, формирующих верхнюю и боковые части среднего мозга. Крыша зрительных долей образована серым веществом - несколькими слоями нервных клеток. Тектум у амфибий является наиболее значимым отделом головного мозга. В зрительных долях находятся полости, являющиеся боковыми ответвлениями мозгового (Сильвиева) водопровода (aquaeductus cerebri (Sylvii) , соединяющего четвёртый мозговой желудочек с третьим.
Дно среднего мозга формируют толстые пучки нервных волокон - ножки большого мозга (cruri cerebri), соединяющие передний мозг с продолговатым и спинным.
Передний мозг (prosencephalon) состоит из промежуточного и конечного мозга, лежащих последовательно.
сверху виден как ромб, острыми углами направленный в стороны.
Части промежуточного мозга лежат вокруг вертикально расположенной широкой щели третьего мозгового желудочка (ventriculus tertius). Боковые утолщения стенок желудочка - зрительные бугры или таламус (thalamus). У рыб и амфибий таламус имеет второстепенное значение (как координационные чувствительные и двигательные центры). Перепончатая крыша третьего мозгового желудочка - надбугорье или эпиталамус - не содержит нейронов. В его составе находится верхняя мозговая железа - эпифиз (epiphisis). У амфибий эпифиз уже выполняет роль железы, но не потерял ещё черты теменного органа зрения. Перед эпифизом промежуточный мозг покрыт перепончатой крышей, орально заворачивающейся внутрь и переходящей в переднее сосудистое сплетение (сосудистую покрышку третьего желудочка), а затем в концевую пластинку промежуточного мозга. Книзу желудочек сужается, формируя воронку гипофиза (infundibulum), каудовентрально к ней крепится нижняя мозговая железа - гипофиз (hypophisis). Впереди, на границе между дном конечного и промежуточного отделов головного мозга находится перекрест зрительных нервов (chiasma nervorum opticorum ). У амфибий большая часть волокон зрительных нервов не задерживается в промежуточном мозге, а идёт дальше - к крыше среднего мозга.
Конечный мозг (telencephalon) по своей длине почти равен длине всех остальных отделов головного мозга. Он состоит из двух частей: обонятельного мозга и двух полушарий, отделённых друг от друга сагиттальной (стреловидной) щелью (fissura sagittalis).
Полушария конечного мозга (haemispherium cerebri) занимают задние две трети конечного мозга и нависают над передней частью промежуточного мозга, частично прикрывая его. Внутри полушарий находятся полости - боковые мозговые желудочки (ventriculi lateralis), каудально сообщающиеся с третьим желудочком. В сером веществе полушарий головного мозга амфибий можно различить три участка: дорсомедиально находится старая кора или гиппокамп (archipallium, s. hippocampus), латерально - древняя кора (paleopallium) и вентролатерально - базальные ядра, соответствующие полосатым телам (corpora striata) млекопитающих. Полосатые тела и, в меньшей степени, гиппокамп являются коррелятивными центрами, последний связан с обонятельной функцией. Древняя кора является исключительно обонятельным анализатором. На вентральной поверхности полушарий заметны борозды, отделяющие полосатые тела от древней коры.
Обонятельный мозг (rhinencephalon) занимает переднюю часть конечного мозга и образует обонятельные доли (луковицы) (lobus olfactorius), спаянные посередине друг с другом. От полушарий они отделяются сбоку краевой ямкой. В обонятельные доли спереди входят обонятельные нервы.
От головного мозга лягушки отходят 10 пар черепномозговых нервов. Их образование, ветвление и зона иннервации принципиально не отличаются от таковых у млекопитающих
Головной мозг птиц.

Ромбовидный мозг (rhombencephalon) включает в себя продолговатый мозг и мозжечок.
Продолговатый мозг (myelencephalon, medulla oblongata) сзади непосредственно переходит в спинной мозг (medulla spinalis). Спереди он вклинивается между зрительными долями среднего мозга. Продолговатый мозг имеет толстое дно, в котором залегают ядра серого вещества - центры многих жизненно важных функций организма (в том числе равновесно-слуховой, соматические двигательные и вегетативные). Серое вещество у птиц покрыто толстым слоем белого, образованного нервными волокнами, соединяющими головной мозг со спинным. В дорсальной части продолговатого мозга имеется ромбовидная ямка (fossa rhomboidea), представляющая собой полость четвертого мозгового желудочка (ventriculus quartus). Крыша четвёртого мозгового желудочка образована перепончатой сосудистой покрышкой, у птиц она полностью прикрыта задним отделом мозжечка.
Мозжечок (cerebellum) у птиц крупный и представлен, практически, только червячком (vermis), расположенным над продолговатым мозгом. Кора (серое вещество, расположенное поверхностно) имеет глубокие борозды, значительно увеличивающие её площадь. Полушария мозжечка развиты слабо. У птиц хорошо развиты отделы мозжечка, связанные с мышечным чувством, в то время как отделы, отвечающие за функциональную связь мозжечка с корой полушарий, практически отсутствуют (они развиваются только у млекопитающих). На продольном разрезе хорошо видны полость желудочка мозжечка (ventriculus cerebelli), а также чередование белого и серого вещества, образующие характерный рисунок древа жизни (arbor vitae).
Средний мозг (mesencephalon) представлен двумя очень крупными, сдвинутыми вбок зрительными долями (lobus opticus s. tectum opticus). У всех позвоночных размер и развитие зрительных долей связан с размером глаз. Они хорошо видны сбоку и с вентральной стороны, тогда как с дорсальной стороны почти полностью прикрыты задними отделами полушарий. В зрительные доли у птиц приходят практически все волокна зрительного нерва, и зрительные доли остаются крайне важными частями головного мозга (однако у птиц со зрительными долями в значимости начинает соперничать кора полушарий). На сагиттальном разрезе видно, что по направлению вперёд полость четвёртого желудочка, суживаясь, переходит в полость среднего мозга - мозговой или силъвиев водопровод (aquaeductus cerebri). Орально водопровод переходит, расширяясь, в полость третьего мозгового желудочка промежуточного мозга. Условная передняя граница среднего мозга образована задней комиссурой (comissura posterior), хорошо заметной на сагиттальном разрезе в виде белого пятнышка.
В составе переднего мозга (prosencephalon) находятся промежуточный и конечный мозг.
Промежуточный мозг (diencephalon) у птиц снаружи виден только с вентральной стороны. Среднюю часть продольного разреза промежуточного мозга занимает узкая вертикальная щель третьего желудочка (ventriculus tertius). В верхней части полости желудочка видно отверстие (парное), ведущее в полость бокового желудочка - монроево (межжелудочковое) отверстие (foramen interventriculare).
Боковые стенки третьего мозгового желудочка образованы достаточно хорошо развитым у птиц таламусом (thalamus), степень развития таламуса связана со степенью развития полушарий. Он, не имея у птиц значения высшего зрительного центра, тем не менее, выполняет важные функции как двигательный коррелятивный центр.
В передней стенке третьего желудочка лежит передняя комиссура (comissura anterior), состоящая из соединяющих два полушария белых волокон
Дно промежуточного мозга именуют гипоталамусом (hypothalamus). При рассмотрении снизу видны боковые утолщения дна - зрительные тракты (tractus opticus). Между ними в передний конец промежуточного мозга входят зрительные нервы (nervus opticus), образующие зрительный перекрест (chiasma opticum). Задний нижний угол третьего мозгового желудочка соответствует полости воронки (infunbulum). Снизу воронка обычно прикрыта хорошо развитой у птиц подмозговой железой - гипофизом (hypophysis).
От крыши промежуточного мозга (эпиталамус (epithalamus) вверх отходит имеющая полость ножка пинеалъного органа. Выше находится и сам пинеалъный орган - эпифиз (epiphysis), он виден сверху, между задним краем больших полушарий и мозжечком. Передняя часть крыши промежуточного мозга образована сосудистым сплетением, заходящим в полость третьего желудочка.
Конечный мозг (telencephalon) у птиц состоит из больших полушарий (hemispherium cerebri), отделенных друг от друга глубокой продольной щелью (fissura interhemispherica). Полушария у птиц являются самыми крупными образованиями головного мозга, но структура их коренным образом отличается от таковой у млекопитающих. В отличие от мозга многих млекопитающих, сильно увеличенные полушария мозга птиц не несут борозд и извилин, поверхность их как с вентральной, так и с дорсальной стороны - гладкая. Кора в целом развита слабо, в первую очередь, в связи с редукцией органа обоняния. Тонкая медиальная стенка полушария переднего мозга в верхней части представлена нервным веществом старой коры (archipallium). Материал новой коры (слабо развита) (neopallium) вместе со значительной массой полосатых тел (corpus striatum) образует толстую боковую стенку полушария или боковой вырост, вдающийся в полость бокового желудочка. Поэтому полость бокового желудочка (ventriculus lateralis) полушария представляет собой узкую щель, расположенную дорсомедиально. У птиц в отличие от млекопитающих в полушариях значительного развития достигает не кора полушарий, а именно полосатые тела. Выявлено, что полосатые тела отвечают за врождённые стереотипные поведенческие реакции, в то время, как новая кора обеспечивает способности к индивидуальному обучению. У птиц некоторых видов обнаружено лучшее, чем в среднем, развитие участка новой коры - это, например, известные своими способностями к обучению вороны.
Обонятельные луковицы (bulbis olfactorius) располагаются на вентральной стороне переднего мозга. Они имеют небольшие размеры и примерно треугольную форму. Спереди в них входит обонятельный нерв.
Земноводные, или амфибии, во взрослом состоянии, как правило, наземные животные, но они еще тесно связаны с водной средой, а их личинки постоянно обитают в воде. Следовательно, русское и греческое (амфибиос - ведущий двойную жизнь) названия отражают главную черту этих позвоночных. Земноводные произошли, как указывалось выше, от девонских кистеперых рыб, обитавших в небольших пресных водоемах и выползавших при помощи своих мясистых парных плавников на берег.
Наружное строение.
Тело (рис. 147) состоит из головы, туловища, передних и задних парных расчлененных конечностей. Конечности состоят из трех отделов: передние - из плеча, предплечья и кисти, задние - из бедра, голени и стопы. Хвост имеется лишь у меньшинства современных земноводных (отряд хвостатые - тритоны, саламандры и др.). Он редуцирован у взрослых форм самой большой группы земноводных - бесхвостых (лягушки, жабы и др.) в связи с приспособлением последних к передвижению прыжками по суше, но сохранился у их личинок - головастиков, обитающих в воде. У немногих видов, ведущих полуподземный образ жизни (отряд безногие, или червяги) редуцировались конечности и хвост.
Голова подвижно сочленяется с туловищем, хотя движение ее очень ограничено и выраженной шеи нет. Расчлененные конечности и подвижное соединение головы с туловищем - характерные признаки наземных позвоночных
, у рыб они отсутствуют. Тело наземных форм сплющено в дорзо-вентраль-ном направлении, в то время как у рыб (в связи с приспособлением их к плаванию) оно, как правило, сдавлено с боков. У водных амфибий форма тела приближается к рыбообразной. Величина тела колеблется от 2 до 160 см (японская саламандра); в среднем амфибии имеют меньшую величину, чем остальные наземные животные. Кожа голая, богатая железами, отделенная во многих местах от мускулатуры благодаря наличию подкожных лимфатических полостей. Она снабжена большим количеством кровеносных сосудов и выполняет также дыхательную функцию (см. далее). У некоторых видов выделения кожных желез ядовиты. Окраска кожи очень разнообразна.
Нервная система.
В связи с приспособлением амфибий к жизни на суше и в особенности в связи с коренным изменением характера передвижения нервная система довольно сильно изменилась. Передний мозг у земноводных (см. рис. 133,В) большей величины, чем средний; у рыб, как правило, наблюдается обратное соотношение. Это объясняется тем, что у рыб функции переднего мозга связаны только с восприятием обонятельных раздражений, у земноводных он начинает принимать участие в координации разных функций тела и в его поверхностном слое появляются зачатки коры (пока еще очень слабые), в которой сосредоточены нервные клетки. В то же время следует отметить, что в переднем мозге хорошо развиты обонятельные доли. Мозжечок у амфибий развит в отличие от рыб очень слабо. Рыбы постоянно движутся, причем положение их тела неустойчивое, земноводные же, опираясь на ноги, находятся в довольно устойчивом положении. Участки спинного мозга, в местах отхождения от него нервов, идущих к мышцам ног, которые выполняют значительно большую работу, чем мышцы парных плавников рыб, утолщены и с ними связаны плечевое и поясничное сплетения нервов. Периферическая нервная система сильно изменилась в связи с дифференциацией мускулатуры (см. дальше) и появлением длинных, членистых конечностей.

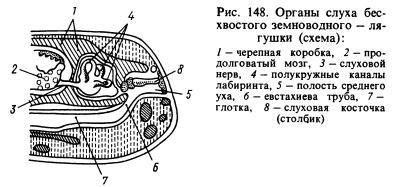
Из органов чувств наиболее существенным изменениям подвергся орган слуха. Передача звуковых волн из водной среды в ткани животного, которые тоже насыщены водой и имеют приблизительно такие же акустические свойства, как и вода, происходит значительно лучше, чем из воздуха. Звуковые волны, распространяющиеся в воздушной среде, почти Целиком отражаются от поверхности животного и только около 1 % энергии этих волн проникает в его тело. В связи с этим у земноводных возник в добавление к лабиринту, или внутреннему уху, новый отдел органа слуха - среднее ухо. Оно представляет собой (рис. 148) наполненную воздухом маленькую полость, сообщающуюся с ротовой полостью посредством евстахиевой трубы и закрытую снаружи тонкой, Упругой барабанной перепонкой. В среднем ухе находится слуховая досточка (или столбик), которая одним концом упирается в барабанную перепонку, а другим - в окошечко, затянутое пленкой и ведущее в полость черепа, где находится лабиринт, окруженный перилимфой. Давление внутри среднего уха равно атмосферному и барабанная перепонка может вибрировать под влиянием звуковых воздушных волн, воздействие которых дальше через слуховую косточку и перилимф передается стенкам лабиринта и воспринимается окончаниями слухового нерва. Полость среднего уха образовалась из первой жаберной щели, а столбик - из расположенной около щели подъязычнонижнечелюстной кости (гиомандибуляре), которая подвешивала висцеральный отдел черепа к мозговому там, где за ушными костями находился лабиринт.
Глаза прикрыты подвижными веками, которые защищают органы зрения от высыхания и засорения. Благодаря изменению формы роговицы и хрусталика земноводные видят дальше, чем рыбы. Амфибии хорошо воспринимают небольшие температурные изменения. Они чувствительны к воздействиям различных веществ, растворенных в воде. Их орган обоняния реагирует на раздражения, вызываемые газообразными веществами. Таким образом, органы чувств земноводных претерпели ряд изменений в связи с переходом к обитанию на суше. У личинок и взрослых животных, обитающих постоянно в воде, имеются, как и у рыб, органы боковой линии.
Земноводным свойственны довольно сложные инстинктивные действия, особенно в период размножения. Например, самец жабы-повитухи, обитающей в России на западе Украины, наматывает на свои задние конечности «шнуры» икры и до развития головастиков прячется в укромных местах на берегу. Через 17-18 дней самец возвращается в воду, где происходит вылупление головастиков. Это своеобразный инстинкт защиты потомства. У ряда тропических бесхвостых амфибий известны еще более сложные инстинкты. Земноводным свойственны и условные рефлексы, однако они вырабатываются с большим трудом.
Двигательная система и скелет. Мышечная система в связи с различными приспособлениями к жизни на суше (развитие конечностей наземного типа, возникновение подвижного сочленения головы с туловищем и др.) подверглась коренным преобразованиям, хотя и сохранила много особенностей, присущих рыбам. Мышечная система рыб очень однообразна и в основном состоит из сходных боковых мышечных сегментов. У земноводных мышечная система стала более дифференцированной, состоящей из разнообразных мышц (рис. 149). У амфибий заложены основы той мышечной системы, которая потом развивалась и усложнялась у настоящих сухопутных позвоночных - пресмыкающихся, птиц и млекопитающих. Это относится и к скелету.
Череп амфибий имеет много хрящевых элементов, что, вероятно, объясняется необходимостью облегчить в связи с полуназемным образом жизни массу тела. В составе черепа много костей, перечисленных при описании черепа высших рыб, и в том числе характерный только для рыб и земноводных парасфеноид (рис. 150). Так как гиомандибуляре стала слуховой косточкой, то роль подвеска выполняет квадратная кость. В связи с утратой во взрослом состоянии жаберного аппарата жаберные дуги редуцируются и сохраняются лишь их видоизмененные остатки. Подъязычная дуга сильно изменяется и частично редуцируется. Череп амфибий очень широкий, что отчасти связано с особенностями их дыхания. Нижняя челюсть, как и у костных рыб, состоит из нескольких костей.
Позвоночный столб (рис. 150) у бесхвостых очень короткий и заканчивается длинной косточкой - уростилем, образовавшимся из рудиментов хвостовых позвонков. У хвостатых земноводных хвостовой отдел позвоночного столба состоит из ряда позвонков. У этих амфибий хвост играет существенную роль при передвижении: в воде он служит для плавания, на суше - для поддержания равновесия. Ребра слабо развиты (у хвостатых земноводных) или редуцированы, а их остатки слились с поперечными отростками позвонков (у остальных амфибий), у древних амфибий ребра имелись. Редукция их у современных форм объясняется необходимостью облегчить массу тела (сильно возросшую при переходе из водной среды в воздушную) у этих позвоночных, еще недостаточно приспособленных к передвижению по суше. Вследствие редукции ребер у земноводных нет грудной клетки. Первый позвонок устроен иначе, чем у рыб: он имеет две суставные впадины для сочленения с двумя затылочными мыщелками черепа, благодаря чему голова земноводных стала подвижной.
Скелет передней конечности (рис. 150) состоит из плечевой кости, двух костей предплечья - лучевой и локтевой, костей запястья, пястных костей и фаланг пальцев. Скелет задней конечности (рис. 150) состоит из бедра, двух костей голени - большой берцовой и малой берцовой, костей предплюсны, плюсневых костей и фаланг пальцев. Следовательно, сходство в строении обеих пар конечностей, несмотря на некоторые различия в их функциях, очень велико. Первоначально передние и задние ноги были пятипалые, у современных земноводных число пальцев может быть меньшим. Задние конечности у многих бесхвостых амфибий служат и для плавания, в связи с чем они удлинились, а пальцы соединены плавательными перепонками.
Пояса конечностей развиты значительно лучше, чем у рыб. Плечевой пояс состоит из костных и хрящевых элементов: лопатки, ключицы, вороньей кости (коракоида) и др. (рис. 150). Ключицы и коракоиды соединены с грудиной, в состав которой тоже входят костные и хрящевые элементы. Головка плеча сочленяется с плечевым поясом. Задний пояс конечностей, или таз, состоит из трех костей: подвздошной, лобковой и седалищной (рис. 150). Образуемая этими костями большая вертлужная впадина служит для сочленения с головкой бедра. Таз соединен с одним позвонком - крестцовым, благодаря чему задние ноги в отличие от брюшных плавников рыб получили довольно прочную опору.
Кровеносная система. У личинок земноводных, обитающих в воде и дышащих жабрами, кровеносная система в основных чертах сходна с кровеносной системой рыб, у взрослых же животных, ведущих наземный образ жизни, она существенно меняется в связи с заменой жаберного дыхания легочным, усилением кожного дыхания, развитием конечностей сухопутного типа и другими изменениями тела. Сердце (см. рис. 134, В, 151) состоит из трех камер: правого и левого предсердий и одного желудочка. От правой части последнего отходит артериальный конус (он был и у рыб, предков земноводных), от которого берут начало четыре пары артерий: первая пара - сонные артерии , несущие кровь в голову, вторая и третья пары - сосуды, соединяющиеся в самый большой сосуд тела - аорту , разветвления которой направляются в разные части тела, четвертая пара - легочнокожные артерии , которые потом разделяются на самостоятельные кожные и легочные артерии.
Из легких кровь, насыщенная кислородом, через легочные вены попадает в левое предсердие, а кровь, насыщенная во всех частях тела углекислым газом, в передней части тела вливается в переднюю полую вену, а в задней части тела - в заднюю полую вену (рис. 152). Обе полые вены впадают в венозный синус , откуда кровь (насыщенная углекислым газом) вливается в правое предсердие. Из обоих предсердий кровь поступает в единственный желудочек сердца. Внутренняя поверхность желудочка имеет углубления и поэтому кровь в нем не успевает полностью смешаться: в левой части его - кровь, насыщенная кислородом, в правой части - кровь, насыщенная углекислым газом, а в средней части - смешанная. Поскольку артериальный конус начинается с правой стороны желудочка, то первая порция крови, попадающая в него (т. е. артериальный конус), будет венозная, она направляется в самые задние артерии - легочные.
Идущая затем смешанная кровь попадает в артерии, образующие аорту, а через разветвления последней во все части тела. Насыщенная кислородом кровь из левой части желудочка направляется в сонные артерии. К этому нужно добавить, что кровь, насыщенная в коже кислородом, попадает, как уже отмечалось выше, через переднюю полую вену и венозный синус в правое предсердие и таким образом разбавляет находящуюся там венозную кровь, далее выталкиваемую в сосуды, образующие аорту. Следовательно, благодаря описанным выше приспособлениям, а также другим, не описанным здесь, разные части тела получают кровь, неодинаково насыщенную кислородом. На рис. 152 изображены главные артериальные и венозные сосуды амфибий.
У земноводных в связи с сильным развитием конечностей и большим, чем у рыб, расчленением тела, существенно изменилась сеть кровеносных сосудов. Появилось много новых сосудов, отсутствовавших у рыб, и сложилась система сосудов, характерная для наземных позвоночных. В то же время следует помнить, что кровеносная система амфибий значительно проще, чем у высших позвоночных.
Дыхательная система. Почти у всех земноводных есть легкие (см. рис. 151; 153). Эти органы имеют еще очень простое строение и представляют собой тонкостенные мешки, в стенках которых разветвляется довольно густая сеть кровеносных сосудов. Так как внутренняя стенка легких почти гладкая, то их поверхность относительно невелика. Трахея почти неразвита и легкие связаны непосредственно с гортанью. Поскольку грудной клетки у земноводных нет (см. выше), акт дыхания обеспечивается работой мышц ротовой полости. Вдох происходит следующим образом. При открытых ноздрях (которые в отличие от ноздрей рыб сквозные, т. е. кроме наружных ноздрей имеются и внутренние ноздри - хоаны ) и закрытом рте оттягивается дно обширной ротовой полости и в нее поступает воздух. Затем ноздри закрываются особыми клапанами, дно ротовой полости поднимается и воздух нагнетается в легкие. Выдох происходит в результате сокращения брюшной мускулатуры.
Значительное количество кислорода земноводные получают через кожу и слизистые оболочки ротовой полости. У некоторых видов саламандр легких совсем нет и весь газообмен совершается через кожу. Однако кожа только в том случае может выполнять функции дыхания, если она влажная. Поэтому обитание земноводных в условиях недостаточной для них влажности воздуха невозможно. Личинки, живущие в воде, дышат жабрами (сначала - наружными, потом - внутренними) и кожей. У некоторых хвостатых земноводных, постоянно живущих в воде, жабры сохраняются всю жизнь. Таким образом, по способам дыхания амфибии еще близки к рыбам.
Выделительная система. Почки (см. рис. 136,А, Б; рис. 151), как и у рыб, туловищные. Вольфовы каналы впадают в клоаку. Туда же открывается мочевой пузырь, в котором накапливается моча. Удаление продуктов диссимиляции происходит, кроме того, через кожу и легкие.
Пищеварительная система. Ротовая полость очень широкая. У ряда видов (главным образом у хвостатых амфибий) имеется много мелких, однородных, примитивно устроенных зубов, которые сидят на челюстях, сошнике, небных и других костях и служат лишь для удержания добычи. У большинства видов (главным образом у бесхвостых амфибий) зубы частично или полностью редуцируются, зато у них сильно развивается язык. Последний у лягушек прикреплен передним концом и может для поимки добычи далеко выбрасываться задним концом вперед. Он очень липкий и хорошо приспособлен для выполнения указанной функции. У видов, постоянно живущих в воде, язык обычно редуцируется. Захват добычи у таких земноводных осуществляется челюстями.
Пищеварительная трубка (см. рис. 151) сравнительно короткая и состоит из глотки, пищевода, желудка, тонкой кишки и совсем небольшой прямой (толстой) кишки. Задняя часть прямой кишки представляет собой клоаку; через нее выводятся кроме кала моча и половые продукты. В ротовую полость впадают слюнные железы, отсутствующие у рыб. Секрет этих желез служит главным образом для смачивания пищи. Слюнные железы очень слабо развиты у видов, обитающих в воде, и значительно лучше - у наземных. Печень велика; поджелудочная железа хорошо выражена. Пища взрослых земноводных в основном животная (насекомые, мелкие позвоночные и др.). Головастики же бесхвостых амфибий большей частью растительноядны.

Размножение. Мужские половые железы (семенники) лежат около почек (см. рис. 151, Б). Их протоки открываются в канальцы передней части почек (см. рис. 136, А) и семя выводится, как и моча, через вольфовы каналы. Женские половые железы (яичники) сильно разрастаются в период икрометания. Икра выходит через очень длинные мюллеровы каналы (см. рис. 136, Б). Последние не имеют прямой связи с яичниками и созревающая икра попадает через полость тела в воронки мюллеровых каналов.
Оплодотворение в большинстве случаев происходит в воде. У многих амфибий этому предшествует сближение самца и самки: самец обхватывает сзади самку, надавливает передними конечностями на ее брюшную стенку и этим способствует выходу в воду икры, которую он сразу оплодотворяет. Таким образом, при наличии полового акта оплодотворение происходит вне организма самки. У меньшинства видов (например, у тритонов) самец выделяет семя в особом мешочке (сперматофоре), который самка тут же захватывает краями клоаки. В данном случае полового акта нет, но оплодотворение - внутреннее. Наконец, у некоторых видов самец вводит семя в клоаку самки при помощи своей способной выпячиваться клоаки.
У многих видов хорошо выражен половой диморфизм (в окраске, строении передних ног, которыми самцы удерживают самок, и по другим признакам). Самцы ряда видов могут издавать очень громкие звуки благодаря усилению этих звуков голосовыми мешками - резонаторами.
Развитие. Развитие земноводных, как правило, происходит в воде. Из оплодотворенных яиц развиваются личинки (головастики), имеющие рыбообразную форму. Дышат они жабрами и их внутреннее строение напоминает рыб. В течение периода роста происходит превращение (метаморфоз) головастиков: сначала у них вырастают задние ноги, потом передние, жабры и хвост атрофируются (у бесхвостых), развиваются легкие, большие изменения происходят в кровеносной системе и т. д.
Происхождение. Земноводные, как было разъяснено выше (с. 296), произошли от кистеперых рыб. Парные плавники древних кистеперых рыб, из которых развились членистые конечности наземных позвоночных, были коротки и широки, в их состав входило множество мелких костных элементов, не соединенных суставами, расположенных в несколько (не менее восьми) поперечных рядов. Пояса, на которые опирались плавники, были сравнительно слабо развиты (особенно тазовый пояс). В связи с превращением плавников в конечности наземного типа произошли существенные изменения в скелете.
Во-первых, редуцировались многие костные элементы: в первых проксимальных рядах осталось по одной кости, в передней ноге - плечо, в задней - бедро; во вторых рядах - по две кости, в передней ноге - лучевая и локтевая кости, в задней - большая и малая берцовые кости; в двух последующих рядах осталось по пять костей, в передней ноге образовавших запястье, в задней - предплюсну; в следующем ряду оставшиеся пять костей вошли в передней ноге в состав пястья, в задней - плюсны; остальные три ряда с пятью костями в каждом стали фалангами пальцев. Уменьшение количества костей способствовало усилению прочности ног.
Во-вторых, кости двух первых рядов (т. е. плеча и предплечья, бедра и голени) сильно удлинились, что было очень важно для усиления быстроты движения.
В-третьих, между перечисленными костями развились суставы, т. е. конечности стали членистыми, что является важнейшим условием их работы.
В-четвертых, укрепились пояса конечностей (см. выше описание поясов). Параллельно указанным изменениям произошли глубокие изменения в нервной, мышечной и сосудистой системах ног. Об изменении других систем органов, происшедших при превращении кистеперых рыб в амфибий, рассказано в общей характеристике последних.
Самыми древними земноводными были стегоцефалы (рис. 154), которые были многочисленны в каменноугольном периоде и окончательно вымерли в триасе. Они жили по берегам водоемов и проводили много времени в воде. Голова этих животных была покрыта щитками, чем и объясняется их название (стегоцефалы - покрытоголовые). В их скелете было много примитивных черт: костные элементы ног были малы и слабо отличались между собой по величине, позвонки двояковогнутые, пояса конечностей плохо развиты и т. д. От них произошли современные группы амфибий.
Мышечная система . Отличается от таковой рыбы главным образом большим развитием мускулатуры конечностей и большей дифференцировкой туловищной мускулатуры, состоящей из сложной системы отдельных мышц. В результате первичная сегментация мускулатуры нарушается, хотя в некоторых брюшных и спинных мышцах она еще остается вполне отчетливой.
Нервная система . Головной мозг земноводных отличается от мозга рыб главным образом большим развитием переднего мозга, полным разделением его полушарий и недоразвитым мозжечком, представляющим собой лишь небольшой валик из нервного вещества, прикрывающего переднюю часть IV желудочка. Развитие переднего мозга выражается не только в его увеличении и дифференцировке, но и в том, что, кроме дна боковых желудочков, их бока и крыша содержат нервное вещество, т. е. у земноводных появляется уже настоящий мозговой свод — архипаллиум (из современных рыб архипаллиум имеется у двоякодышащих). Обонятельные доли лишь слабо отграничены от полушарий. Промежуточный мозг сверху лишь слегка прикрыт соседними отделами. К крыше его прикрепляется теменной орган, а от дна отходит хорошо выраженная воронка, к которой прикрепляется гипофиз. Средний мозг хотя и представляет собой значительный отдел, но он относительно меньше, чем у рыб. Недоразвитие мозжечка, как и у двоякодышащих рыб, связано с несложностью движений тела: земноводные— вообще малоподвижные животные, у тех же из них, которые подобно лягушкам могут совершать быстрые движения, они ограничиваются прыжками, т. е. движениями очень простыми. От головного мозга, как и у костистых рыб, отходят только 10 пар головных нервов; XII пара (подъязычный нерв) отходит за пределами черепной коробки, а XI пара (добавочный нерв) вообще не развита.

. I — сверху; II — снизу; III — сбоку; IV — в продольном разрезе (по Паркеру):
1 — полушария переднего мозга, 2 — обонятельная доля, 3 — обонятельный нерв, 4 — промежуточный мозг, 5 — зрительная хиазма, 6 — воронка, 7 — гипофиз, 8 — средний мозг, 9 — мозжечок, 10 — продолговатый мозг, 11 — четвертый желудочек, 12 — спинной мозг, 13 — третий желудочек, 14 — сильвиев водопровод,
III —X — головные нервы, XII — подъязычный нерв

, схема (по Грегори):
1 — черепная коробка, 2 — продолговатый мозг, 3 — слуховой нерв, 4 — полукружные каналы, 5 — полость среднего уха, 6 — евстахиева труба, 7 — глотка, 8 — етремя, 9 — барабанная перепонка
Настоящих спинномозговых нервов у лягушки 10 пар. Три передние пары принимают участие в образовании плечевого сплетения, иннервирующего передние конечности, а четыре задние пары—в образовании пояснично-крестцового сплетения, иннервирующего задние конечности.
Симпатическая нервная система лягушки, как у всех земноводных, развита очень хорошо и представлена главным образом двумя нервными стволами, которые тянутся с обеих сторон позвоночника и образованы цепью нервных узлов, связанных друг с другом тяжами и соединенных со спинномозговыми нервами.
Еще интересные статьи






